Multiple Inflammatory Signaling Cascades in Inflammatory Breast Cancer, Taking into Account Chemoresistance and Recurrence of the Tumor and Type 2 Diabetes Mellitus. (Audit of Differences in Carcinogenic Pathways)
by Oleksii Volodimirovich Movchan1, Irina Yuriivna Bagmut2*, Ivan Ivanovich Smolanka1, Igor Leonidovich Kolisnyk2, Oleksandr Vasyliovych Bagmut3, Andriy Oleksandrovich Lyashenko1, Irina Viktorivna Dosenko1, Oksana Mykolaivna Ivankova1, Anton Dmitrovich Loboda1
1Department of Breast Cancer and Reconstructive Surgery, National Cancer Institute of Ukraine of Ministry of Health of Ukraine
²Department of Clinical Pathophysiology, Topographic Anatomy and Operative Surgery, Kharkiv National Medical University of the Ministry of Health of Ukraine, Kharkiv, Ukraine
3Faculty of Computer Sciences, Karazin Kharkiv National University, Ukraine
*Corresponding author: Irina Yuriivna Bagmut, Department of Clinical Pathophysiology, Topographic Anatomy and Operative Surgery, Department of Kharkiv National Medical University of the Ministry of Health of Ukraine, Kharkiv, Ukraine
Received Date: 15 May 2025
Accepted Date: 21 May 2025
Published Date: 23 May 2025
Citation: Movchan OV, Bagmut IY, Smolanka II, Kolisnyk IL, Bagmut OV, et al. (2025) Multiple Inflammatory Signaling Cascades in Inflammatory Breast Cancer, Taking into Account Chemoresistance and Recurrence of the Tumor and Type 2 Diabetes Mellitus. (Audit of Differences in Carcinogenic Pathways). J Surg 10: 11329 https://doi.org/10.29011/2575-9760.011329
Abstract
Introduction: Advanced Breast Cancer (ABC) with type 2 diabetes mellitus could be an uncommon, however profoundly forceful frame of breast cancer, which accounts for less than 5% of all locally progressed introductions.
The Aim of the Study: aimed to current understanding of breast tumor chemoresistant - breast cancers. Localized at essential breast area and treated early can still relapse since of the presence of cancer cells and the change of cancer cells into a moderately forceful phenotype.
Methods: We determined the the clinical introduction of advanced breast cancer regularly contrasts altogether from that of non-advanced breast cancer; immunohistochemistry uncovers few recognizing highlights. The more forceful triple-negative and HER2-positive breast cancer subtypes are overrepresented in advanced breast cancer compared to non- advanced breast cancer, with a poorer guess in reaction to routine treatments and plasma glucose level control with hyperglucosemia modification.
Results: The part of the tumor microenvironment, through tumor-associated macrophages, infiltrating lymphocytes is additionally examined, recommending that these tumors outward variables may offer assistance account for the contrasts in behavior between advanced breast cancer with high plasma glucose level and non- advanced breast cancer.
Conclusions: This audit summarizes the current prove recommending that inflammatory signaling pathways are up regulated in this illness, which may give a road for novel therapeutics against the background of diabetes. There are different novel treatment techniques as of now underway in clinical trials; they require for encourage the improvement of preclinical models of this uncommon but forceful illness is vital.
Keywords: Advanced breast cancer; Cancer chemoresistant and Relapse; Diabetes mellitus; Legal and Ethical relations; Signaling pathways
Introduction
Breast cancer remains a dangerous disease, indeed with all the recent technological headways. In time, the performed intercession has made an affect, but an overwhelmingly huge number of breast cancer patients still live beneath the fear of chemoresistant and “recurrent” disease [1]. Over the long time, a few variables have been examined with the overarching point of being able to prognoses chemoresistant disease. Current understanding of breast tumor chemoresistant - breast cancers localized at essential breast area and treated early can still relapse since of the presence of cancer cells and change of cancer cells into a moderately forceful phenotype [2]. A tall rate of forceful cells ordinarily checks chemoresistant and recurrent breast cancers. Breast cancer survivors are at hazard for creating unused cancers for a few reasons-whatever caused unique cancer might still have an impact, either on moment primaries within the same organ or on related cancers in other organs [3], i.e. are relatively non-specific for this type of cancer, such as Type 2 Diabetes Mellitus (T2DM) [4]. Although many efforts have been made, molecular or pathological diagnostic criteria for ABC have not yet been identified. The molecular subtypes of ABC patients include hormone receptor-positive (HR+, positive for estrogen and/or progesterone receptors)/HER2+ (14.8%), HR+/HER2-negative (HER2−; 35.7%), HR−/HER2+ (23.1%), and triple-negative (TN-IBC, negative for estrogen and progesterone receptors and HER2; 26.4%) [5]. Among these ABC subtypes, TN-ABC carries the worst prognosis [6]. The standard treatment for ABC is trimodality therapy, which consists of chemotherapy, surgery, and radiation. Although this therapeutic approach has significantly improved patient survival, the median survival of ABC patients remains poor. Over the past two decades, researchers have identified some molecular changes that play important roles in ABC. These include loss of WNT1-inducible-signaling Pathway Protein 3 (WISP3) [7]; overexpression of epidermal growth factor receptor (EGFR), HER2, TIG1/Axl, E-cadherin, overexpression of inflammatory mediators Janus Kinase (JAK)/Signal Transducers And Activators of Transcription (STAT), nuclear factor kappa B (NF-κB), and Cyclooxygenase-2 (COX-2) [8], but glucose metabolism and DNA repair is observed in approximately 20-30% of malignant neoplasms of this organ and correlates with the aggressive course of the disease [9]. The Tumor Microenvironment (TME), which includes T cells, Tumor-Associated Macrophages (TAMs) showed critical roles in promoting ABC aggressiveness. TAMs enhance ABC cell migration. It has been shown that the presence of tumor-infiltrating lymphocytes was increased in the tumors of ABC patients who had a pathological complete response (pCR) to neoadjuvant chemotherapy [10]. But, one of the main criteria was plasma glucose level control, because it can lead to adverse event like hyperglycemia above 14.0 mmol/l same way hypoglycemia below 4.0 mmol/l [11]. When to summarize critical signaling pathways in the progression and possible chemoresistant and relapse of ABC, crosstalk with the ABC TME, and the up-to-date preclinical and clinical studies of targeting these pathways in ABC (Figure 1).
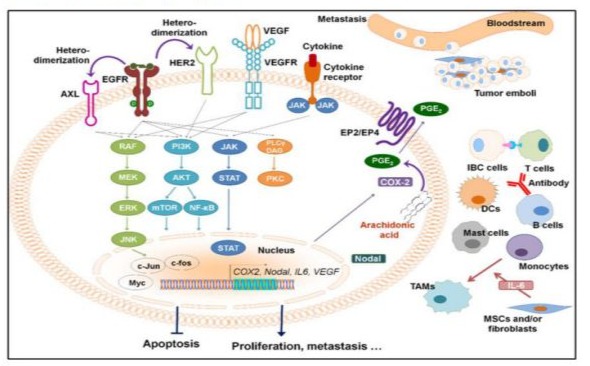
Figure 1: Overexpressed or activated signaling pathways in inflammatory breast cancer and the tumor microenvironment. The presence of tumor emboli, in which cells pack together to form a tumor cell cluster, is a hallmark of IBC cells. DCs, dendritic cells; ABC, advanced breast cancer; MSCs, mesenchymal stem cells; TAMs, tumor-associated macrophages. [Wang X, Semba T, Phi LTH, et al. Targeting Signaling Pathways in Inflammatory Breast Cancer. Cancers (Basel). 2020;12(9):2479. Published 2020 Sep 1. doi:10.3390/cancers12092479].
Body of Review
Molecular Characteristic of ABC: Accordingly, the major molecular subtypes described for non-ABC also exist within ABC, with the associated prognostic and histological features. The most frequently mutated genes were TP53 and PIK3CA, with alterations spanning the whole coding sequence of TP53 and involving hotspot regions in PIK3CA. Moreover, recurrent copy number gains were identified in MYC, CCND1, FGF19, FGF3, and FGFR1. Among them, CCND1, FGF19, and FGF3 genes mapped on the same cytogenetic band, 11q13.3, and showed a statistically significant co-occurrence (p value < 0.001 and q < 0.01). In particular, TP53, PIK3R1, and NF1 mutations were detected more frequently in triple negative tumors (odds ratio > 2.71, p value < 0.05). CCND1, FGF3, and FGFR1 copy number gains were identified in luminal cases (odds ratio < 0.36, p value < 0.05). The diversity of genomic alterations occurring within ABC patients, indicating an opportunity for more personalized therapies that specifically target oncoproteins encoded by mutated or amplified genes [12].
Inflammation and ABC: Excessive abundance of cytokines and chemokines in the tumor microenvironment are well-recognized factors that underpin the progression of solid malignancies [13]. These soluble factors not only support survival, proliferation, and invasion of tumor cells but also increase angiogenesis and facilitate the evasion of immune surveillance. A comprehensive study Morrow and colleagues [14] of 36 surgical ABC samples after neoadjuvant therapy analyzed the expression of 538 genes implicated in tumor-associated inflammation and angiogenesis. Authors failed to detect any significant differences in the expression of many inflammatory cytokines and chemokines, including IFN-γ, TNF, IL-1α, IL-1β, IL-8, and IL-10 between ABC and non-ABC tissue samples, indicating that the associated inflammatory phenotype commonly visible is more likely due to blockage of the dermal lymphatics by disseminated tumor cells than by an infiltration of inflammatory cells [15]. The latent transcription factor components of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) and signal transducer and activator of transcription- (STAT-) 3, with a lesser involvement of the RAS/mitogen-activated protein kinase (MAPK)/Jun kinase (JNK) pathway and cyclooxygenase (COX) enzymes. Indeed, emerging evidence suggests that these pathways may also play an important role in ABC [16].
Inflammatory Signaling Pathways
NF-κB Pathway and Associated Cytokines: The NF-κB pathway induces the expression of a variety of pro-inflammatory cytokines and chemokines and regulates inflammasomes. IL-6 and IL-8 are two cytokines regulated by the NF-κB pathway and are hyperactivated in ABC. SUM149 and SUM190 cells secreted significant amounts of both IL-6 and IL-8 compared with non-ABC cell lines. Inhibition of the NF-κB pathway by using tetrathiomolybdate, a specific copper chelator, or by transfecting a dominant-negative IκBα vector reduced the secretion of VEGF, fibroblast growth factor 2 (FGF2), IL-1α, IL-6, and IL-8 in vitro and SUM149 tumor growth and angiogenesis in vivo. The NF-κB family consists of P65, RelB, cRel, P105, and P100, which require dimerization in order to elicit transcriptional activity [17]. The activation of NF-κB signaling can occur via classical and alternative pathways. Classical NF-κB pathway activation occurs in response to inflammatory stimuli, including TNF, IL-1β, and activators of toll-like receptors, while alternative NF-κB activation occurs in response to ligand engagement of members of the TNF receptor superfamily, such as RANK, Fn14, lymphotoxin β receptor, and CD40. Сlassical NF-κB pathway modulates expression of genes involved in cell proliferation, survival, innate immunity, inflammation, and angiogenesis, while genes regulated by the alternative NF-κB pathway regulate homeostasis of adaptive immunity and lymphangiogenesis [18]. In a cancer setting, tumors are often characterised by elevated levels of cytokines produced and secreted by the classical pathway, including two proinflammatory cytokines, TNF and IL-1β, which commonly trigger its activation. Accordingly, constitutive activation of NF-κB, which results in upregulation of antiapoptotic proteins, is frequently observed in ER-negative/HER2-positive tumors [19]. Fliegauf and colleagues [20] - the linkage interval was robust to various assumptions about whether individuals with milder phenotypes should be considered as “affected.” Although the coding exons of several genes in the candidate region, including P100 (which spans ∼116 kbp and comprises 23 coding exons plus a non-coding first exon, results might be attributed to an underestimation of sequence variants adjacent to the coding exons, such as the non-coding intronic splice sites, which were not exhaustively studied at that time. Using the SUM-149 IBC cell line, it was observed that classical and alternative NF-κB pathway activity both promoted the formation of tumorspheres in vitro, suggesting that these pathways may regulate the function of tumor-initiating cells [21]. Furthermore, the inflammatory cytokines IL-6 and IL-8, which are among the best characterized NF-κB target genes, are produced and secreted at high levels within ABC [22]. Оverexpression of IL-1β has been identified within the serum of patients with ER-negative breast tumors. Enhanced cell motility and invasion have also been coupled with overexpression of IL-1β within breast cancer via the upregulation of matrix metalloproteinase- 9, integrin-1. The secretion of IL-1β requires inflammasome activation and processing of pro IL-1β by Caspase-1 or 8. Inflammasome signaling becomes activated upon pathogen or danger-associated molecular patterns, or activation of programmed cell death machinery [23].
JAK/STAT Pathway and Associated Cytokines: The JAK/STAT pathway contributes to many cellular processes, including immunity, cell growth, cell death, and differentiation. STAT activation is associated with the formation of various cancers, including melanoma, prostate cancer, non-ABC, and ABC. Researchers led by samples from ABC patients were enriched in CD44+/CD24− CSCs and activated phosphor-STAT3 (pSTAT3) cells, and 40% of CD44+/CD24− cells were positive for pSTAT3, suggesting STAT pathway activation in ABC CSCs. They also demonstrated that the inhibition of JAK2 reduced the proliferation and tumor growth of pSTAT3+ ABC cells in vitro and in vivo [24]. Therefore, the researchers further showed that inhibiting the recruitment of TAMs decreased pSTAT3 expression and IL-6 secretion within the TME, leading to delayed ABC tumor formation and reduced local recurrence [25]. These results demonstrated that IL-6 mediates the tumor-promoting influences of the ABC TME. Importantly, phosphorylated STAT3 (pSTAT3) is particularly abundant on the leading edge of tumors, as well as in surrounding lymphocytes and stromal cells, suggesting a role in invasion and metastasis. Constitutive activation of STAT3 has also shown to accelerate tumor progression and increase the metastatic potential in HER2-positive breast cancers. A retrospective study conducted by Di Bonito M. et colleagues [6] - serum IL-6 concentrations are not only increased in more than half of all breast cancer patients, but are also significantly higher in ABC patients compared to non-ABC patients. Similar to other solid malignancies, elevated IL-6 expression in breast cancer positively correlates with increased tumor stage, lymph node involvement, recurrence risk. Observed, that failure of HER2 and other-directed therapies coincides with IL-6-mediated activation of STAT3. IL-6-dependent activation of the JAK2/STAT3 has shown promise as a therapeutic target for hormone receptor-negative and HER2-positive breast cancers, as these subtypes produce higher levels of IL-6 [26]. This signaling cascade was found to be prevalent for the viability of the ABC SUM-190 cell line [27].
COX Pathway
Levels of COX enzymes are increased within 40% of breast cancers, while prostaglandin E2, the main product catalysed by COX-2, is produced at high levels in various human breast cancer cell lines. Expression of COX-2 has been confirmed in 13 breast cancer cell lines by qRT-PCR, with no detection in normal breast tissue [28]. COX-2 expression also correlates with poor prognostic indicators, such as increased tumor size, axillary node and distant metastasis, tumor grade, high-proliferation rates, receptor-negative disease, and HER2 amplification [29]. The molecular ABC signature defined by Fernandez and colleagues [10] suggested elevated COX-2 expression in IBC compared to non-ABC tumors, and this was also reflected by more abundant prostaglandin E2 in primary ABC tumors. ABC cell lines also have higher levels of PGE2 and PGF2α, two enzymatic products of COX-2, than do non-IBC cell lines. High COX-2 expression correlates with worse OS and higher nuclear grade in ABC patients. In addition, the COX-2 pathway contributes to invasiveness and the CSC population in ABC. As a key inflammatory molecule in ABC, COX-2 is functionally linked to other signaling pathways. It is worth noting that the COX-2 pathway has emerging roles in modulating the tumor immune microenvironment. COX-2-derived PGE2 has been shown to mediate the crosstalk between colonic tumor cells and macrophages [30].
Tumor-Associated Macrophages/Monocytes
Tumor-associated macrophages (TAMs) produce and secrete high levels of inflammatory mediators, that not only promote survival and proliferation of neoplastic cells but also antagonize the antitumor activity of CD8-positive T cells. Wolfe and colleagues [31] described the accumulation of CD68-positive macrophages in the normal tissue surrounding ABC lesions. TAMs also contribute to the chemoresistant and relapses of ABC cells via releasing mediators of invasion and angiogenesis including TNF, IL-6, IL-8, and IL-10. The expression of these cytokines is significantly higher in CD14-positive tumor-infiltrating monocytes of ABC patients than in those from non-ABC patients [32].
Tumor-Infiltrating Lymphocytes
The presence of tumor-infiltrating lymphocytes and in particular, the proportion of functional cytotoxic CD8-positive T cells have been suggested to predict patient response to immune checkpoint treatment. In contrast, the presence of exhausted T cells with poor effector function is typically associated with the expression of programmed death-ligand 1 (PD-L1) and other immune checkpoint inhibitors on tumor cells and/or tumor-infiltrating lymphocytes. Thus, high PD-L1 expression may negatively regulate T cells, thus preventing the activation and migration of CD8-positive T cells into ABC tumors [33]. IHC staining identified aggregates of CD8-positive T cells as major subpopulations associated with intratumoral and peritumoral desmoplastic stroma in approximately half of ABC tumors analysed, with low density of single-spread cells across other samples [34]. However, these tumors stained minimally for the regulatory T cell marker FoxP3, while tumor-associated staining of the immune checkpoint regulator PD-L1 varied greatly. Not surprisingly, the same authors observed a positive correlation between the extent of CD8-positive T cell infiltration and mutation rate as a predictor of variability of neoantigen, which in turn correlated with the presence of several mutations in gene-encoding DNA mismatch repair genes [35].
ABC and Diabetes Mellitus
Advanced breast cancer, the p53 gene mutation is observed mainly in tumors with plasma glucose level above 10.0 mmol/l. The data obtained indicate that mutations in the p53 gene are observed with a significantly higher frequency in the presence of microsatellite instability in the loci of chromosomes responsible for post-replication repair. In these tumors, despite the preserved ability to restore chromosomal breakdowns, the emergence and further development of the tumor are associated with another mechanism of carcinogenesis – mutations in the "genome guard" p53 gene – with a subsequent conformational change in the quaternary structure of the protein and loss of its control function, and subsequent violation of apoptosis. It is known that the rate of tumor growth is one of the most important integral indicators of the features of its clinical course. It should be understood that the growth rate is determined by the balance between the activity of proliferative processes with plasma glucose level (is an aggravating factor) on the one hand, and apoptosis on the other [36].
Discussion
Narod et al. [37] correctly identify diagnosis—defined as “the probability that a tumor if left unattended, would not become clinically apparent or cause death” and largely a consequence of screening—as a relevant issue. This review is an attempt to check for differences in the carcinogenic pathways of ABC (the multimutational model of carcinogenesis) to determine the level of disease aggression and possible ways to target cancer cells of ABC patients by influencing specific pathways. Thus, we understand the diversity of developmental forms of ABC, in which inflammatory factors (multiple inflammatory signaling cascades have been identified as deregulated and overactive within ABC lesions, contributing to the creation of inflammatory loops between cancer cells of ABC and the cells of the surrounding tumor microenvironment). No full controls received chemotherapy. The risk of chemoresistant to breast cancer and recurrence development was approximately the same. It is likely that biomarkers will be identified in the future that can better predict cancer chemoresistant and recurrence, but this possibility does not negate the validity of the present observations.
Conclusions and Perspective
The cause of breast cancer chemoresistant and recurrence and possible strategies to prevent it remains elusive. There is an urgent need to decipher the complex relationship between the individual components that lead to relapse, but also to devise effective therapies with minimal toxicity. Talking about complex interrelationships, there is enough evidence to link Molecular Characterization of ABC (the various molecular determinants of tumor recurrence are very intricately connected and interregulated, making it difficult to establish a hierarchical sense and plasma glucose level), Inflammation and ABC, NF-κB Pathway and Associated Cytokines, JAK/STAT Pathway and Associated Cytokines, COX Pathway, Tumor-Associated Macrophages/Monocytes, Tumor-Infiltrating Lymphocytes, plasma glucose level above 10.0 mmol/l. All these hot spots of cellular / physiological events were observed in chemoresistant and recurrent breast cancers. Although individual reports have verified the existence of one or more of these, it is essential to note that we are far from determining the causal reason for breast cancer chemoresistant and recurrence. With the intricate relationship between these individual factors, it will always be difficult to guess which one comes first. A factor that has severely hindered our progress in this field is the absence of an acceptable model for the study of tumor relapses. For a meaningful study, it is crucial to promote healthy collaboration between basic scientists and clinical researchers. Breast cancer chemoresistant and recurrence is too complex a problem to be fully understood only through laboratory investigations or clinical observations. Essential to our understanding of ABC as a disease is the creation of further preclinical models, which will allow us to more confidently explore and confirm whether inflammatory processes are indeed contributing to the progression of ABC more than in an ABC-free environment. The presence of MSI is somehow associated with hyperglucosemia and is an aggravating factor in the course of advanced breast cancer with a poorer response to specific therapy. The wealth of literature that has accumulated in recent years is a good start. These observations suggest that, in the context of chemoresistant and relapses, they are as advanced as their invasive counterpart. We tried to look for and find a much higher overall risk of breast cancer chemoresistant and recurrence after an ABC diagnosis.
References
- Aguilera T, Rafat M, Castellini L, Shehade H, Kariolis M (2016) Reprogramming the immunological microenvironment through radiation and targeting Axl. Nat. Commun 7: 13898.
- Preda M, Ilina R, Potre O, Potre C, Mazilu O (2020) Survival Analysis of Patients with Inflammatory Breast Cancer in Relation to Clinical and Histopathological Characteristics. Cancer Manag Res 12: 12447-12455.
- Ueno N, Fernandez J, Cristofanilli M, Overmoyer B, Rea D (2018) International Consensus on the Clinical Management of Inflammatory Breast Cancer from the Morgan Welch Inflammatory Breast Cancer Research Program 10th Anniversary Conference. J. Cancer 9: 1437-1447.
- Miricescu D, Totan A, Stanescu-Spinu II, Badoiu SC, Stefani C, et al. (2020) PI3K/AKT/mTOR Signaling Pathway in Breast Cancer: From Molecular Landscape to Clinical Aspects. Int J Mol Sci 22: 168-173.
- Wang X, Semba T, Phi L (2020) Targeting Signaling Pathways in Inflammatory Breast Cancer. Cancers (Basel) 12: 2479-2487.
- Di Bonito M, Cantile M, Botti G (2019) Pathological and molecular characteristics of inflammatory breast cancer. Translational Cancer Research 8: 449-456.
- Liu Yi (2019) The emerging role of WISP proteins in tumorigenesis and cancer therapy. Journal of Translational Medicine 17: 28.
- Morrow R, Etemadi N, Yeo B, Ernst M (2017) Challenging a Misnomer? The Role of Inflammatory Pathways in Inflammatory Breast Cancer. Mediators Inflamm 2017: 4754827.
- Movchan O, Bagmut I (2021) Impacts of Inflammatory Microenvironment on the Development of Cancer Relapse after Combined Treatment. Ukraïnsʹkij žurnal medicini, bìologìï ta sportu 6: 204-213.
- Fernandez SV, MacFarlane AW, Jillab M (2020) Immune phenotype of patients with stage IV metastatic inflammatory breast cancer. Breast Cancer Res 22: 134-137.
- Amrith BP, Jain P, Chaudhari KA, Sharma M, Batra U (2019) Severe hypoglycemia with trastuzumab: An unseen adverse event. South Asian J Cancer 8: 140-149.
- Fumagalli C, Ranghiero A, Gandini S (2020) Inter-tumor genomic heterogeneity of breast cancers: comprehensive genomic profile of primary early breast cancers and relapses. Breast Cancer Res 22: 107-110.
- Gallagher A (2020) A 26-Year Study Highlights Invasive Breast Cancer Recurrence After treatment. Oncology Times 42: 28-35.
- Mussbacher M, Salzmann M, Brostjan C, Hoesel B, Schoergenhofer C (2019) Cell Type-Specific Roles of NF-κB Linking Inflammation and Thrombosis. Front Immunol 10: 85-95.
- Giannakeas V, Sopik V, Narod S (2018) Association of radiotherapy with survival in women treated for ductal carcinoma in situ with lumpectomy or mastectomy. JAMA Network Open 1: e181100.
- Yan S, Vandewalle N, De Beule N, Faict S, Maes K (2019) AXL Receptor Tyrosine Kinase as a Therapeutic Target in Hematological Malignancies: Focus on Multiple Myeloma. Cancers (Basel) 11: 1727-1738.
- Yang P, Peng Q, Lian W, Fu F, Wang C (2022) Validation of the Eighth American Joint Committee on Cancer Anatomic and Prognostic Staging System for Breast Cancer. Journal of Surgical Research 270: 539-546.
- Zhou Y, Cui C, Ma X, Luo W, Zheng S (2020) Nuclear Factor κB (NF-κB)-Mediated Inflammation in Multiple Sclerosis. Front Immunol 11: 391-399.
- Czauderna C (2019) “Context-Dependent Role of NF-κB Signaling in Primary Liver Cancer-from Tumor Development to Therapeutic Implications. Cancers (Basel) 11: 1053-1061.
- Fliegauf M, Bryant VL, Frede N (2015) Haploinsufficiency of the NF-κB1 Subunit p50 in Common Variable Immunodeficiency. Am J Hum Genet 97: 389-403.
- Lei M, Lee T (2021) Cancer Stem Cells: Emerging Key Players in Immune Evasion of Cancers. Front Cell Dev Biol 9: 692940.
- Lee J, Liu H, Pearson T, Iwase T, Fuson J (2021) PI3K and MAPK Pathways as Targets for Combination with the Pan-HER Irreversible Inhibitor Neratinib in HER2-Positive Breast Cancer and TNBC by Kinome RNAi Screening. Biomedicines 9: 740.
- Acevedo-Díaz A, Morales-Cabán B, Zayas-Santiago A, Martínez-Montemayor M, Suárez-Arroyo I (2022) SCAMP3 Regulates EGFR and Promotes Proliferation and Migration of Triple-Negative Breast Cancer Cells through the Modulation of AKT, ERK, and STAT3 Signaling Pathways. Cancers 14: 2807-2817.
- Ruxolitinib W (2014) Preop Chemo For Triple Negative Inflammatory Brca 2014.
- Hibino S, Kawazoe T, Kasahara H, Itoh S, Ishimoto T (2021) Inflammation-Induced Tumorigenesis and Metastasis. International Journal of Molecular Sciences 22: 5421-5429.
- Hidalgo-Estévez M, Stamatakis K, Jiménez-Martínez M, López-Pérez R, Fresno M (2020) Cyclooxygenase 2-Regulated Genes an Alternative Avenue to the Development of New Therapeutic Drugs for Colorectal Cancer. Frontiers in Pharmacology 11: 112-116.
- Quail DF, Joyce JA (2013) Microenvironmental regulation of tumor progression and metastasis. Nat. Med 19: 1423-1437.
- Reyes M (2014) Targeting COX-2 and Rank in aggressive Breast Cancers: Inflammanory Breast Cancer and Triple-Negative Breast Cancer. DISSERTATION for the Degree of Doctor of Philosophy 2014.
- Wolfe A, Trenton N, Debeb B, Larson R, Ruffell B (2016) Mesenchymal stem cells and macrophages interact through IL-6 to promote inflammatory breast cancer in pre-clinical models. Oncotarget 7: 82482-82492.
- Shihab I, Khalil B, Elemam N, Hachim I, Hachim M (2020) Understanding the Role of Innate Immune Cells and Identifying Genes in Breast Cancer Microenvironment. Cancers 12: 2226-2235.
- Jiao D, Zhang J, Zhu J (2021) Comparison of survival in non-metastatic inflammatory and other T4 breast cancers: a SEER population-based analysis. BMC Cancer 21: 138-148.
- Johnson A, Kleczko E, Nemenoff R (2020) Eicosanoids in Cancer: New Roles in Immunoregulation. Front Pharmacol 11: 595-598.
- Heeren A, Rotman J, Stam A, Pocorni N, Gassama A (2019) Efficacy of PD-1 blockade in cervical cancer is related to a CD8+FoxP3+CD25+ T-cell subset with operational effector functions despite high immune checkpoint levels. J Immunother Cancer 7: 43-49.
- Matsuda N, Wang X, Lim B, Krishnamurthy S, Alvarez R (2018) Safety and Efficacy of Panitumumab Plus Neoadjuvant Chemotherapy in Patients with Primary HER2-Negative Inflammatory Breast Cancer. JAMA Oncol 4: 1207-1213.
- David Y, Serena S, Siddharth S, Tony L, McCarthy E (2020) Intratumoral CD4+ T Cells Mediate Anti-tumor Cytotoxicity in Human Bladder Cancer, Cell 181: 1612-1625.
- Bagmut I, Movchan O, Sheremet M, Smolanka I, Lyashenko A, et al. (2022) Characteristics of certain genetical and biological properties of carcinogenesis in the development of inflammatory breast cancer with type 2 diabetes mellitus and tumor relapse. Rom J Diabetes Nutr Metab Dis 29: 245-252.
- Narod S, Rakovitch E (2014) A comparison of the risks of in-breast recurrence after a diagnosis of dcis or early invasive breast cancer. Curr Oncol 21: 119-124.
© by the Authors & Gavin Publishers. This is an Open Access Journal Article Published Under Attribution-Share Alike CC BY-SA: Creative Commons Attribution-Share Alike 4.0 International License. Read More About Open Access Policy.